iModel
Related models
DS_Gabor_One
MEO_Gabor
ME_Gabor
MEO_Gabor_Rot
ME_Gabor_Rot
RD_Exp_T
RD_2Gabor
RD_2Gabor_Rect
Variations
MEO_Gabor.sHtQ
MEO_Gabor.sHtH
MEO_Gabor.sHt1
MEO_Gabor.sHt2
MEO_Gabor.s1tQ
MEO_Gabor.s1tH
MEO_Gabor
MEO_Gabor.s1t2
MEO_Gabor.s2tQ
MEO_Gabor.s2tH
MEO_Gabor.s2t1
MEO_Gabor.s2t2
Opponent Motion Energy, Gabor Filter
- Adelson EH, Bergen JR (1985) Spatiotemporal energy models
for the perception of motion. J Opt Soc Am A 2:284-299.
- Bair W, Movshon JA (2004) Adaptive temporal integration of
motion in direction-selective cells in macaque visual cortex. J
Neurosci 24:7305--7323.
- Grzywacz NM, Yuille AL (1990) A model for the estimate of
local image velocity by cells in the visual cortex. Proc Roy Soc Lond
B 239:129-161.
- van Santen JPH, Sperling G (1984) Temporal covariance model of
human motion perception. J Opt Soc Am A 1:451--473.
- van Santen JPH, Sperling G (1985) Elaborated Reichardt detectors. J Opt Soc Am A 2:300--321.
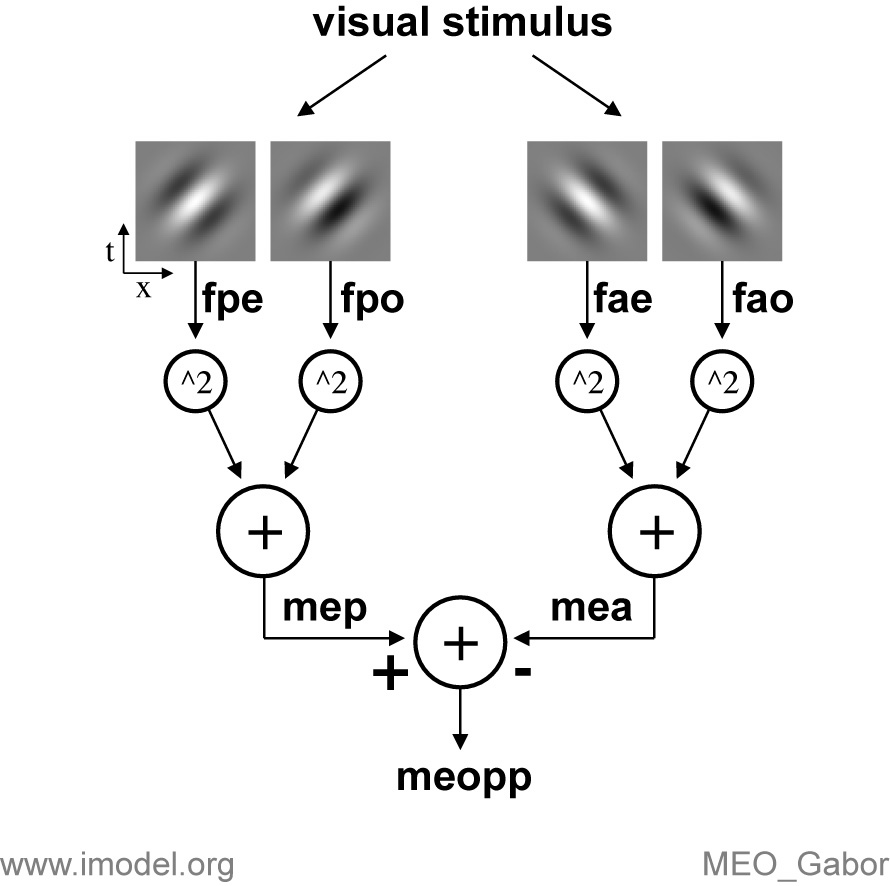
The visual stimulus is processed (convolved) by four linear Gabor
filters (icons show x-t slices of 3D filters) to yield the signals
fpe (filter preferred even), fpo (filter preferred odd),
fae (anti-preferred even) and fao (anti-preferred odd).
Each filter output is squared, and the signals are added for each
quadrature pair to give the motion energy in the preferred direction,
mep, and the anti-preferred direction, mea. These two
oppositely tuned ME signals are subtracted to give the opponent motion
energy, meopp.
The opponent signal (meopp) is then offset, scaled and half-wave rectified, and it is used to drive a Poisson spiking mechanism. The spikes are time shifted to simulate a neurobiological latency. See the model (.moo) files for the parameters that govern these computations.